Circadian rhythm refers to endogenous biological rhythm, which completes cyclic repetitions every 24 hours; the term originated from the Latin words circa, which means “around,” and dies, which means “day” [1,2]. Most organisms on the planet including human beings have evolved their own rhythmic programming of biological and behavioral outputs in response to these stimuli to maintain metabolic balance and promote survival [3,4]. Circadian rhythm helps our physiology to adapt to the environmental or behavioral changes in a time-of-day-dependent manner [5]. Beyond simply adapting to stimulus-response interaction, circadian rhythms are governed by an intrinsic molecular oscillator, the circadian clock [6]. Zeitgeber, literally meaning “time giver,” refers to any stimulus related to time cue from the external environment [1,6]. Our circadian clock constantly communicates with these zeitgebers to maintain circadian rhythm originally settled [1-6]. Without external stimulation, circadian clocks quite predictably control the daily timing of synthesis and release of most hormones [1,7].
In mammals, the circadian clock affects many aspects of the physiology including sleep-wake cycles, feeding-fasting cycles, and metabolic processes of the gastrointestinal tract, kidney, heart, and liver [8,9]. Previous epidemiological studies demonstrated that specific diseases such as myocardial infarction, hypertensive crisis, and asthma attacks occur at certain times during the day, supporting the relationship between circadian clock and human pathophysiology [10,11]. Moreover, rapidly developing social conditions such as artificial lighting, 24 h accessibility to fast food, and temperature control by advanced heating and cooling system have threatened this powerful and ancient circadian clock [5]. A growing population of shift workers and travelers across time zones experience fatigue, disorientation, and insomnia because of disruption of circadian rhythm [12]. Thus, exceeding the adaptive limitations of circadian clock leads to negative effects of circadian control on metabolic balance, which can be explained by a dramatic increase in the rates of obesity, diabetes, and so on [12]. Therefore, we need to understand the role of circadian control in metabolism because this can be a novel clue for the treatment of metabolic syndrome.
Since Nobel Prize in Physiology or Medicine was awarded in 2017, research on the circadian clock and its inner workings for metabolism has been actively continued [13]. For a long time, the concept of time-restricted feeding (TRF) for the enhanced treatment of diseases such as obesity and metabolic syndrome has highlighted the influence of the circadian clock on metabolic regulation [14,15]. This review will summarize how the circadian clock affects metabolism by identifying its components involved in metabolic syndrome and their implications.
COMPONENTS OF CIRCADIAN CLOCK
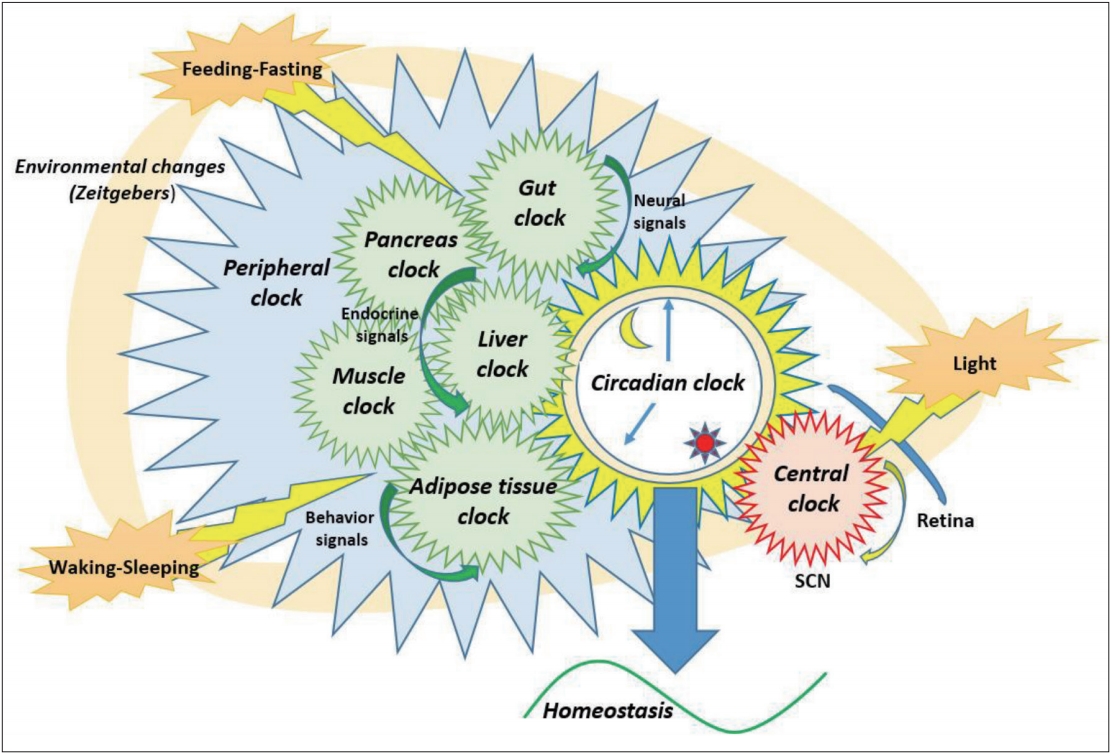
Circadian clock has a hierarchical architecture; it is composed of two major classes: the central master clock in the hypothalamic superchiasmatic nucleus (SCN) and the peripheral clock within non-SCN cells distributed in various organs such as the gut, muscle tissue, liver, white adipose tissue, brown adipose tissue, and pancreas (Figure 1) [9,12]. The SCN functions as the master controller and perceives light information, which is one of the most important zeitgeber for resetting the central pacemaker, via the retina and transmits neuronal or circulating humoral signals to peripheral clocks to generate circadian rhythms in the whole body [5,12].
This circadian timing system at the molecular level is dependent on the transcriptional-translational feedback loops formed by changing mRNA expression, protein stability, and metabolite production [5]. The two core genes are circadian locomotor output cycles kaput (CLOCK) and brain and muscle-Arnt-like 1 (BMAL1) [5,16]. These transcriptional activators, CLOCK and BMAL1, heterodimerize each other and bind to enhancer sequences to rhythmically induce transcription of genes, including the period genes (PER) and the cryptochrome genes (CRY), for the negative feedback loop. Expression of RAR-orphan receptors (ROR α and γ) induced by CLOCK-BMAL1 activates transcription for circadian rhythm. Casein kinase I δ and ε (CKI δ and ε) also contribute to the activation of the clock function via destabilization and degradation of PER proteins. FBXL3 degrades CRY proteins, functioning as an activator. On the contrary, besides PER and CRY, reverse erythroblastosis virus (Rev-erbs) suppress BMAL1 at the transcriptional level (Table 1). These molecular oscillators exist in all cells of the body [5,11].
CIRCADIAN CONTROL OF METABOLIC HOMEOSTASIS–THE SIX CLOCKS
The regulation of metabolism by circadian clock is reciprocal and varies in amplitude and phase according to endocrine factors and organs involved; however, it usually shows 24 h periodicity following the earth’s rotation [2]. The master clock (the SCN) controls the peripheral clocks by sending neuronal (via the autonomic nervous system), hormonal, and behavioral signals and by changing body temperature [2,8,11]. However, glucose ingestion rather than light is a more potent zeitgeber for peripheral tissues when it comes to circadian control of metabolic process [10]. The circadian machinery controlling glucose metabolism consists of six clocks: the central clock (the SCN), gut clock, muscle clock, adipose tissue clock, liver clock, and pancreatic clock (Table 2) [9].
The central clock directly controls glucose uptake, because food intake happens in a wake cycle, affecting insulin sensitivity of the entire body [4]. The SCN plays an indirect role by releasing several hormones related to glucose tolerance. Insulin secretion is reduced by cortisol, melatonin, and growth hormone. These endocrine hormones are known to show daily oscillations governed by the hypothalamic-pituitary axes [1]. The gut clock affects intestinal motility and nutrient absorption by the rhythmic expression of membrane glucose transporters and brush border disaccharides [17]. The human skeletal muscles have an autonomous molecular clock regulating muscle-insulin sensitivity and mitochondrial respiration [18,19]. Circadian control of lipid mobilization, that is lipolysis and lipogenesis, is mediated by the adipose tissue clock [20,21]. Adipose tissue also produces adipokines including leptin and adiponectin contributing to endocrine signaling responsible for appetite cycle [20,21]. The liver center, referred to as the fuel management center, regulates gluconeogenesis in addition to mitochondrial dynamics [22]. Lastly, pancreatic islets are involved in insulin biosynthesis and transport and glucose-stimulated insulin secretion, contributing to the diurnal rhythm of insulin and glucagon secretion [23,24].
CIRCADIAN DISRUPTION AND METABOLIC OUTCOMES
Circadian disruption of metabolism is an emerging risk factor for the metabolic syndrome [25]. This can be classified into two aspects according to zeitgebers: light, associated with sleep and wake cycle, and nutrients, associated with feeding and fasting cycle.
First, disturbed sleep disrupts the circadian rhythm by modulating light exposure, resulting in impaired insulin sensitivity [26]. Sleep deprivation has been associated with a decrease in resting metabolic rate and an increase in postprandial plasma glucose concentrations, which result from circadian misalignment [27]. It also alters levels of hormones; it reduces leptin and increases ghrelin levels, promoting the feeling of hunger during sleep [28]. Shift workers typically experience dyssynchrony of light, sleep, and eating [29]. Accumulating evidence from epidemiological studies shows that shift workers have an increased risk of cardiovascular disease, obesity, diabetes, and other metabolic disorders [30-32]. Chronic jet lag, another example of sleep-wake cycle disruption, has led to the development of leptin resistance and increased the incidence rate of metabolic disorders [33,34]. The possible mechanisms for the effects of disturbed sleep on insulin resistance are changes in sympatho-vagal balance and increasing circulation levels of hormones such as catecholamines and cortisol [35,36].
Second, negative effects of mistimed feeding, that is feeding unexpectedly during the resting period, has been associated with metabolic health concerns [37,38]. This is a well-proven fact based on many experiments with mice. Light-fed mice, which fed during the usual rest period, eat more calories and gain more body mass than night-fed ones. Even though light-fed mice were limited to the same quantity of food, they still gain more body mass, which results from decreased energy expenditure. However, rearrangement of food intake during their original feeding period reversed the metabolic consequences of mistimed eating [39,40]. In humans, skipping breakfast increases the risks of metabolic syndromes including obesity [40,41]. Besides, repeated late dinner at night is strongly associated with increased adiposity and elevated body mass [42,43]. Similarly, TRF can help improve metabolic health by altering clock function [14]. According to Moro et al., the TRF group showed significantly lower fat mass and leptin, triglyceride, insulin growth factor-1, and interleukin-1ß levels and higher adiponectin levels than the normal diet group [44]. Another prediabetes trial demonstrated that early TRF has a beneficial effect on cardiometabolic health because of improved insulin sensitivity, blood pressure, and oxidative stress. [45]. These results show that feeding time is critical for energy balance based on circadian rhythm.
In conclusion, peeking into metabolic pathways via circadian clocks can provide not only an understanding of the mechanisms mediating the metabolic disorders but also the potential therapeutic strategies to restore clock-controlled functions. Furthermore, the zeitgeber that exerts a higher influence on metabolism, between light and feeding, has not been determined yet. Further research on circadian-clock regulation with regard to metabolism and zeitgebers is required to control metabolic complications.